

Electron Micrograph Of Cell Labeled
A typical beast prison cell is x–twenty μm in bore, which is well-nigh one-5th the size of the smallest particle visible to the naked centre. It was non until good light microscopes became bachelor in the early part of the nineteenth century that all establish and animal tissues were discovered to be aggregates of individual cells. This discovery, proposed as the prison cell doctrine by Schleiden and Schwann in 1838, marks the formal birth of prison cell biology.
Fauna cells are non only tiny, they are likewise colorless and translucent. Consequently, the discovery of their main internal features depended on the development, in the latter part of the nineteenth century, of a variety of stains that provided sufficient dissimilarity to make those features visible. Similarly, the introduction of the far more powerful electron microscope in the early 1940s required the development of new techniques for preserving and staining cells before the full complexities of their internal fine structure could begin to emerge. To this day, microscopy depends as much on techniques for preparing the specimen as on the performance of the microscope itself. In the discussions that follow, we therefore consider both instruments and specimen training, beginning with the light microscope.
Effigy 9-one shows a series of images illustrating an imaginary progression from a thumb to a cluster of atoms. Each successive paradigm represents a tenfold increase in magnification. The naked eye could see features in the first two panels, the resolution of the low-cal microscope would extend to about the 4th panel, and the electron microscope to virtually the seventh panel. Some of the landmarks in the development of calorie-free microscopy are outlined in Table ix-1. Figure 9-two shows the sizes of diverse cellular and subcellular structures and the ranges of size that different types of microscopes can visualize.

Figure 9-one
A sense of scale between living cells and atoms. Each diagram shows an image magnified by a cistron of ten in an imaginary progression from a pollex, through peel cells, to a ribosome, to a cluster of atoms forming part of one of the many poly peptide molecules (more...)
Table 9-1
Some Important Discoveries in the History of Light Microscopy.

Figure 9-2
Resolving power. Sizes of cells and their components are drawn on a logarithmic scale, indicating the range of objects that can be readily resolved by the naked eye and in the light and electron microscopes. The following units of length are normally (more than...)
The Light Microscope Can Resolve Details 0.2 μm Autonomously
In general, a given blazon of radiation cannot be used to probe structural details much smaller than its own wavelength. This is a fundamental limitation of all microscopes. The ultimate limit to the resolution of a calorie-free microscope is therefore gear up by the wavelength of visible light, which ranges from about 0.4 μm (for violet) to 0.7 μm (for deep reddish). In applied terms, bacteria and mitochondria, which are almost 500 nm (0.5 μm) wide, are generally the smallest objects whose shape tin be conspicuously discerned in the calorie-free microscope; details smaller than this are obscured past effects resulting from the wave nature of light. To understand why this occurs, nosotros must follow what happens to a beam of light waves equally it passes through the lenses of a microscope (Figure 9-three).
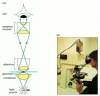
Figure nine-3
A light microscope. (A) Diagram showing the light path in a compound microscope. Light is focused on the specimen by lenses in the condensor. A combination of objective lenses and eyepiece lenses are arranged to focus an paradigm of the illuminated specimen (more than...)
Because of its wave nature, lite does not follow exactly the arcadian straight ray paths predicted by geometrical optics. Instead, light waves travel through an optical system past a variety of slightly different routes, and then that they interfere with 1 another and crusade optical diffraction effects. If two trains of waves reaching the same point by different paths are precisely in stage, with crest matching crest and trough matching trough, they will reinforce each other and then as to increase effulgence. In contrast, if the trains of waves are out of phase, they will interfere with each other in such a way as to abolish each other partly or entirely (Effigy 9-4). The interaction of light with an object changes the phase relationships of the low-cal waves in a way that produces complex interference effects. At loftier magnification, for example, the shadow of a directly border that is evenly illuminated with low-cal of compatible wavelength appears every bit a set of parallel lines, whereas that of a circular spot appears as a set of concentric rings (Effigy ix-v). For the same reason, a unmarried point seen through a microscope appears as a blurred disc, and two betoken objects close together give overlapping images and may merge into one. No amount of refinement of the lenses can overcome this limitation imposed by the wavelike nature of light.

Figure nine-4
Interference between lite waves. When two low-cal waves combine in phase, the aamplitude of the resultant wave is larger and the brightness is increased. 2 calorie-free waves that are out of phase cancel each other partly and produce a wave whose aamplitude, (more than...)

Figure 9-5
Edge effects. The interference furnishings observed at high magnification when light passes the edges of a solid object placed between the light source and the observer are shown here.
The limiting separation at which two objects tin still be seen equally distinct—the so-called limit of resolution—depends on both the wavelength of the light and the numerical aperture of the lens system used. This latter quantity is a measure out of the width of the entry pupil of the microscope, scaled according to its distance from the object; the wider the microscope opens its eye, so to speak, the more than sharply it tin can come across (Figure 9-six). Under the best weather, with violet light (wavelength = 0.4 μm) and a numerical aperture of i.4, a limit of resolution of just under 0.ii μm can theoretically be obtained in the light microscope. This resolution was achieved by microscope makers at the finish of the nineteenth century and is only rarely matched in contemporary, factory-produced microscopes. Although it is possible to enlarge an epitome every bit much as ane wants—for instance, by projecting it onto a screen—it is never possible to resolve two objects in the light microscope that are separated by less than nigh 0.2 μm; they will appear as a single object.

Figure nine-6
Numerical aperture. The path of light rays passing through a transparent specimen in a microscope illustrate the concept of numerical discontinuity and its relation to the limit of resolution.
We see next how interference and diffraction can be exploited to study unstained cells in the living state. Later nosotros hash out how permanent preparations of cells are fabricated for viewing in the light microscope and how chemic stains are used to heighten the visibility of the prison cell structures in such preparations.
Living Cells Are Seen Clearly in a Stage-Contrast or a Differential-Interference-Contrast Microscope
The possibility that some components of the prison cell may be lost or distorted during specimen preparation has e'er challenged microscopists. The only certain mode to avoid the problem is to examine cells while they are alive, without fixing or freezing. For this purpose, light microscopes with special optical systems are specially useful.
When lite passes through a living cell, the phase of the light wave is changed according to the cell'due south refractive index: light passing through a relatively thick or dense part of the cell, such as the nucleus, is retarded; its phase, consequently, is shifted relative to calorie-free that has passed through an adjacent thinner region of the cytoplasm. The phase-contrast microscope and, in a more complex fashion, the differential-interference-contrast microscope, exploit the interference effects produced when these two sets of waves recombine, thereby creating an image of the cell's construction (Figure 9-7). Both types of light microscopy are widely used to visualize living cells.

Effigy 9-seven
Two ways to obtain contrast in light microscopy. (A) The stained portions of the cell reduce the amplitude of light waves of particular wavelengths passing through them. A colored image of the jail cell is thereby obtained that is visible in the ordinary fashion. (more than...)
A simpler manner to meet some of the features of a living jail cell is to observe the light that is scattered by its various components. In the nighttime-field microscope, the illuminating rays of light are directed from the side so that only scattered light enters the microscope lenses. Consequently, the jail cell appears as a vivid object against a nighttime groundwork. With a normal vivid-field microscope, the image is obtained by the simple transmission of light through a cell in civilisation. Images of the same cell obtained by 4 kinds of lite microscopy are shown in Figure nine-8.

Effigy ix-8
Iv types of light microscopy. Four images are shown of the same fibroblast cell in culture. All 4 types of images can exist obtained with most mod microscopes by interchanging optical components. (A) Bright-field microscopy. (B) Phase-contrast microscopy. (more than...)
Phase-contrast, differential-interference-dissimilarity, and dark-field micros-copy go far possible to watch the movements involved in such processes as mitosis and cell migration. Since many cellular motions are too wearisome to be seen in real time, it is often helpful to have time-lapse motion pictures or video recordings. Here, successive frames separated past a short time delay are recorded, then that when the resulting picture series or videotape is played at normal speed, events announced greatly speeded upwards.
Images Can Be Enhanced and Analyzed by Electronic Techniques
In recent years electronic imaging systems and the associated applied science of image processing have had a major impact on light microscopy. They have enabled certain applied limitations of microscopes (due to imperfections in the optical system) to exist largely overcome. They have also circumvented 2 fundamental limitations of the human eye: the eye cannot see well in extremely dim light, and it cannot perceive minor differences in light intensity against a bright background. The first limitation can exist overcome by attaching highly sensitive video cameras (the kind used in dark surveillance) to a microscope. Information technology is so possible to find cells for long periods at very depression light levels, thereby avoiding the damaging effects of prolonged vivid light (and estrus). Such low-calorie-free cameras are particularly important for viewing fluorescent molecules in living cells, as explained beneath.
Considering images produced past video cameras are in electronic class, they tin can be readily digitized, fed to a calculator, and processed in various ways to extract latent information. Such image processing makes information technology possible to recoup for diverse optical faults in microscopes to attain the theoretical limit of resolution. Moreover, by electronic image processing, contrast can be greatly enhanced so that the eye's limitations in detecting small differences in light intensity are overcome. Although this processing likewise enhances the effects of random groundwork irregularities in the optical system, such defects tin can be removed past electronically subtracting an epitome of a blank area of the field. Small transparent objects that were previously impossible to distinguish from the background and then become visible.
The high contrast accessible by computer-assisted differential-interference-contrast microscopy makes it possible to encounter even very modest objects such as single microtubules (Figure 9-9), which have a diameter of 0.025 μm, less than one-10th the wavelength of light. Individual microtubules can also be seen in a fluorescence microscope if they are fluorescently labeled (run into Effigy nine-15). In both cases, however, the unavoidable diffraction effects badly mistiness the image so that the microtubules announced at least 0.ii μm broad, making it impossible to distinguish a single microtubule from a parcel of several microtubules.

Figure 9-9
Paradigm processing. (A) Unstained microtubules are shown here in an unprocessed digital prototype, captured using differential-interference-dissimilarity microscopy. (B) The prototype has now been processed, start by digitally subtracting the unevenly illuminated background, (more than...)
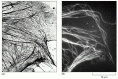
Figure 9-xv
Immunofluorescence. (A) A manual electron micrograph of the periphery of a cultured epithelial jail cell showing the distribution of microtubules and other filaments. (B) The aforementioned expanse stained with fluorescent antibodies confronting tubulin, the protein (more...)
Tissues Are Usually Fixed and Sectioned for Microscopy
To make a permanent preparation that can be stained and viewed at leisure in the microscope, one offset must care for cells with a fixative so as to immobilize, kill, and preserve them. In chemic terms, fixation makes cells permeable to staining reagents and cross-links their macromolecules then that they are stabilized and locked in position. Some of the primeval fixation procedures involved immersion in acids or in organic solvents, such equally alcohol. Current procedures usually include treatment with reactive aldehydes, particularly formaldehyde and glutaraldehyde, which form covalent bonds with the free amino groups of proteins and thereby cross-link adjacent protein molecules.
Most tissue samples are besides thick for their individual cells to be examined directly at high resolution. Afterward fixation, therefore, the tissues are usually cut into very thin slices, or sections, with a microtome, a machine with a sharp bract that operates similar a meat slicer (Figure ix-10). The sections (typically one–ten μm thick) are and then laid apartment on the surface of a glass microscope slide.

Figure 9-10
Making tissue sections. This illustration shows how an embedded tissue is sectioned with a microtome in grooming for examination in the lite microscope.
Considering tissues are more often than not soft and fragile, fifty-fifty after fixation, they need to exist embedded in a supporting medium before sectioning. The usual embedding media are waxes or resins. In liquid form these media both permeate and environs the stock-still tissue; they can then be hardened (past cooling or past polymerization) to form a solid cake, which is readily sectioned by the microtome.
There is a serious danger that whatsoever treatment used for fixation and embedding may alter the construction of the cell or its constituent molecules in undesirable ways. Rapid freezing provides an alternative method of preparation that to some extent avoids this trouble by eliminating the need for fixation and embedding. The frozen tissue can be cut direct with a special microtome that is maintained in a cold bedroom. Although frozen sections produced in this way avoid some artifacts, they suffer from others: the native structures of individual molecules such every bit proteins are well preserved, but the fine structure of the cell is frequently disrupted by ice crystals.
Once sections take been cut, past whatever method, the next footstep is normally to stain them.
Dissimilar Components of the Cell Can Be Selectively Stained
In that location is little in the contents of most cells (which are 70% water by weight) to impede the passage of calorie-free rays. Thus, about cells in their natural state, fifty-fifty if fixed and sectioned, are almost invisible in an ordinary light microscope. One way to brand them visible is to stain them with dyes.
In the early nineteenth century, the demand for dyes to stain textiles led to a fertile flow for organic chemistry. Some of the dyes were found to stain biological tissues and, unexpectedly, often showed a preference for item parts of the cell—the nucleus or mitochondria, for example—making these internal structures clearly visible. Today a rich diverseness of organic dyes is available, with such colorful names as Malachite green, Sudan blackness, and Coomassie bluish, each of which has some specific affinity for particular subcellular components. The dye hematoxylin, for case, has an affinity for negatively charged molecules and therefore reveals the distribution of Dna, RNA, and acidic proteins in a jail cell (Figure nine-11). The chemic basis for the specificity of many dyes, however, is not known.
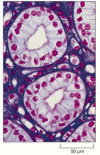
Figure 9-11
A stained tissue section. . This section of cells in the urine-collecting ducts of the kidney was stained with a combination of dyes, hematoxylin and eosin, usually used in histology. Each duct is made of closely packed cells (with nuclei stained ruby) (more...)
The relative lack of specificity of these dyes at the molecular level has stimulated the design of more rational and selective staining procedures and, in particular, of methods that reveal specific proteins or other macromolecules in cells. It is a trouble, however, to accomplish adequate sensitivity for this purpose. Since relatively few copies of about macromolecules are nowadays in any given cell, one or two molecules of stain leap to each macromolecule are often invisible. One way to solve this problem is to increment the number of stain molecules associated with a unmarried macromolecule. Thus, some enzymes can be located in cells through their catalytic activeness: when supplied with appropriate substrate molecules, each enzyme molecule generates many molecules of a localized, visible reaction product. An alternative and much more generally applicable approach to the problem of sensitivity depends on using dyes that are fluorescent, as we explain next.
Specific Molecules Tin Be Located in Cells by Fluorescence Microscopy
Fluorescent molecules blot light at i wavelength and emit it at another, longer wavelength. If such a compound is illuminated at its absorbing wavelength and so viewed through a filter that allows only light of the emitted wavelength to pass, it is seen to glow against a dark background. Because the background is dark, even a infinitesimal amount of the glowing fluorescent dye tin can be detected. The aforementioned number of molecules of an ordinary stain viewed conventionally would be practically invisible because they would give simply the faintest tinge of color to the light transmitted through this stained part of the specimen.
The fluorescent dyes used for staining cells are detected by a fluorescence microscope. This microscope is like to an ordinary calorie-free microscope except that the illuminating calorie-free, from a very powerful source, is passed through two sets of filters—one to filter the low-cal before it reaches the specimen and ane to filter the calorie-free obtained from the specimen. The start filter is selected so that it passes only the wavelengths that excite the item fluorescent dye, while the second filter blocks out this light and passes but those wavelengths emitted when the dye fluoresces (Figure 9-12).

Figure 9-12
The optical arrangement of a fluorescence microscope. A filter set consists of ii barrier filters (1 and three) and a dichroic (beam-splitting) mirror (2). In this example, the filter ready for detection of the fluorescent molecule fluorescein is shown. High-numerical-aperture (more...)
Fluorescence microscopy is well-nigh often used to detect specific proteins or other molecules in cells and tissues. A very powerful and widely used technique is to couple fluorescent dyes to antibody molecules, which and then serve as highly specific and versatile staining reagents that bind selectively to the particular macromolecules they recognize in cells or in the extracellular matrix. 2 fluorescent dyes that have been commonly used for this purpose are fluorescein, which emits an intense green fluorescence when excited with blueish lite, and rhodamine, which emits a deep ruby-red fluorescence when excited with greenish-xanthous light (Figure nine-13). Past coupling i antibody to fluorescein and another to rhodamine, the distributions of different molecules can be compared in the aforementioned prison cell; the two molecules are visualized separately in the microscope by switching back and forth between two sets of filters, each specific for ane dye. As shown in Figure 9-14, three fluorescent dyes tin can be used in the same way to distinguish between three types of molecules in the same cell. Many newer fluorescent dyes, such as Cy3, Cy5, and the Alexa dyes, take been specifically adult for fluorescence microscopy (meet Figure 9-13).

Figure 9-13
Fluorescent dyes. The maximum excitation and emission wavelengths of several normally used fluorescent dyes are shown in relation to the corresponding colours of the spectrum. The photon emitted past a dye molecule is necessarily of lower energy (longer (more...)

Figure ix-14
Multiple-fluorescent-probe microscopy. In this composite micrograph of a cell in mitosis, three different fluorescent probes take been used to stain three dissimilar cellular components. The spindle microtubules are revealed with a green fluorescent antibody, (more...)
Important methods, discussed later on in the chapter, enable fluorescence microscopy to exist used to monitor changes in the concentration and location of specific molecules inside living cells (see p. 574).
Antibodies Tin Be Used to Detect Specific Molecules
Antibodies are proteins produced by the vertebrate immune arrangement as a defence force confronting infection (discussed in Affiliate 24). They are unique among proteins because they are made in billions of different forms, each with a unlike bounden site that recognizes a specific target molecule (or antigen). The precise antigen specificity of antibodies makes them powerful tools for the cell biologist. When labeled with fluorescent dyes, they are invaluable for locating specific molecules in cells by fluorescence microscopy (Figure nine-15); labeled with electron-dense particles such as colloidal gold spheres, they are used for similar purposes in the electron microscope (discussed below).
The sensitivity of antibodies as probes for detecting and assaying specific molecules in cells and tissues is frequently enhanced by chemical methods that dilate the betoken. For example, although a marker molecule such equally a fluorescent dye can be linked directly to an antibody used for specific recognition—the master antibody—a stronger signal is achieved by using an unlabeled primary antibiotic and then detecting it with a grouping of labeled secondary antibodies that demark to it (Figure 9-xvi).

Figure 9-16
Indirect immuno-cytochemistry. This detection method is very sensitive because the primary antibody is itself recognized by many molecules of the secondary antibody. The secondary antibody is covalently coupled to a marker molecule that makes it readily (more...)
The most sensitive amplification methods apply an enzyme as a marker molecule fastened to the secondary antibody. The enzyme alkaline phosphatase, for case, in the presence of appropriate chemicals, produces inorganic phosphate and leads to the local germination of a colored precipitate. This reveals the location of the secondary antibody that is coupled to the enzyme and hence the location of the antibody-antigen circuitous to which the secondary antibody is bound. Since each enzyme molecule acts catalytically to generate many thousands of molecules of product, even tiny amounts of antigen can be detected. An enzyme-linked immunosorbent assay (ELISA) based on this principle is frequently used in medicine every bit a sensitive test—for pregnancy or for various types of infections, for example. Although the enzyme amplification makes enzyme-linked methods very sensitive, improvidence of the colored precipitate away from the enzyme means that the spatial resolution of this method for microscopy may be limited, and fluorescent labels are normally used for the most precise optical localization.
Antibodies are made nigh simply by injecting a sample of the antigen several times into an animate being such as a rabbit or a caprine animal and then collecting the antibody-rich serum. This antiserum contains a heterogeneous mixture of antibodies, each produced by a different antibiotic-secreting cell (a B lymphocyte). The unlike antibodies recognize various parts of the antigen molecule (called an antigenic determinant, or epitope), as well as impurities in the antigen preparation. The specificity of an antiserum for a item antigen can sometimes be sharpened by removing the unwanted antibody molecules that demark to other molecules; an antiserum produced against protein Ten, for instance, can be passed through an affinity column of antigens Y and Z to remove whatever contaminating anti-Y and anti-Z antibodies. Even and so, the heterogeneity of such antisera sometimes limits their usefulness. This problem is largely overcome by the employ of monoclonal antibodies (see Figure eight-six). Even so, monoclonal antibodies can likewise have bug. Since they are unmarried antibiotic poly peptide species, they bear witness near perfect specificity for a single site or epitope on the antigen, just the accessibility of the epitope, and thus the usefulness of the antibody, may depend on the specimen preparation. For example, some monoclonal antibodies volition react only with unfixed antigens, others only after the use of detail fixatives, and still others only with proteins denatured on SDS polyacrylamide gels, and not with the proteins in their native conformation.
Imaging of Complex Three-dimensional Objects Is Possible with the Optical Microscope
For ordinary light microscopy, as we have seen, a tissue has to exist sliced into sparse sections to be examined; the thinner the section, the crisper the image. In the process of sectioning, data almost the third dimension is lost. How, then, can one become a picture of the 3-dimensional architecture of a jail cell or tissue, and how can i view the microscopic structure of a specimen that, for one reason or another, cannot get-go be sliced into sections? Although an optical microscope is focused on a particular focal plane within complex iii-dimensional specimens, all the other parts of the specimen above and below the airplane of focus are also illuminated, and the light originating from these regions contributes to the image as "out-of-focus" mistiness. This tin can brand information technology very hard to interpret the prototype in detail, and can pb to fine image construction being obscured past the out-of-focus light.
Two approaches have been developed to solve this problem: one is computational, the other is optical. These three-dimensional microscopic imaging methods make information technology possible to focus on a chosen aeroplane in a thick specimen while rejecting the lite that comes from out-of-focus regions above and below that airplane. Thus one sees a well-baked, thin optical section. From a series of such optical sections taken at unlike depths and stored in a estimator, it is easy to reconstruct a three-dimensional epitome. The methods practice for the microscopist what the CT scanner does (past different means) for the radiologist investigating a human torso: both machines give detailed exclusive views of the interior of an intact structure.
The computational approach is often called epitome deconvolution. To sympathise how it works, remember how the wave nature of light means that the microscope lens system gives a small blurred disc as the paradigm of a point low-cal source, with increased blurring if the betoken source lies above or below the focal aeroplane. This blurred paradigm of a point source is chosen the indicate spread function. An image of a circuitous object can and then be thought of equally existence congenital up by replacing each point of the specimen by a corresponding blurred disc, resulting in an image that is blurred overall. For deconvolution, we get-go obtain a series of (blurred) images, focusing the microscope in turn on a series of focal planes—in effect, a blurred three-dimensional image. The stack of images is then processed past computer to remove every bit much of the blur as possible. Substantially the computer program uses the microscope'south betoken spread function to determine what the effect of the blurring would take been on the prototype, and and so applies an equivalent "deblurring" (deconvolution), turning the blurred iii-dimensional paradigm into a serial of clean optical sections. The computation required is quite complex, and used to be a serious limitation. All the same, with faster and cheaper computers, the image deconvolution method is gaining in power and popularity. An example is shown in Figure 9-17.

Figure 9-17
Epitome deconvolution. (A) A light micrograph of the large polytene chromosomes from Drosophila, stained with a fluorescent DNA-binding dye. (B) The same field of view after paradigm deconvolution clearly reveals the banding blueprint on the chromosomes. Each (more...)
The Confocal Microscope Produces Optical Sections by Excluding Out-of-Focus Light
The confocal microscope achieves a result similar to that of deconvolution, but does so by manipulation of the light earlier it is measured; thus it is an analog technique rather than a digital ane. The optical details of the confocal microscope are complex, only the basic idea is simple, as illustrated in Figure 9-18.
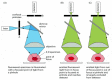
Effigy nine-18
The confocal fluorescence microscope. This simplified diagram shows that the basic organization of optical components is similar to that of the standard fluorescence microscope shown in Figure 9-12, except that a laser is used to illuminate a small pinhole (more than...)
The microscope is by and large used with fluorescence optics (see Figure 9-12), but instead of illuminating the whole specimen at once, in the usual way, the optical system at any instant focuses a spot of calorie-free onto a single signal at a specific depth in the specimen. A very bright source of pinpoint illumination is required; this is usually supplied by a laser whose light has been passed through a pinhole. The fluorescence emitted from the illuminated material is collected and brought to an paradigm at a suitable light detector. A pinhole aperture is placed in front of the detector, at a position that is confocal with the illuminating pinhole—that is, precisely where the rays emitted from the illuminated point in the specimen come up to a focus. Thus, the light from this point in the specimen converges on this aperture and enters the detector.
By contrast, the low-cal from regions out of the plane of focus of the spotlight is likewise out of focus at the pinhole aperture and is therefore largely excluded from the detector (Effigy ix-19). To build up a ii-dimensional paradigm, data from each signal in the plane of focus are nerveless sequentially past scanning across the field in a raster pattern (as on a television screen) and are displayed on a video screen. Although not shown in Figure 9-18, the scanning is ordinarily washed by deflecting the beam with an oscillating mirror placed betwixt the dichroic mirror and the objective lens in such a style that the illuminating spotlight and the confocal pinhole at the detector remain strictly in register.
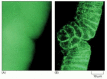
Figure 9-19
Conventional and confocal fluorescence microscopy compared. These two micrographs are of the aforementioned intact gastrula-phase Drosophila embryo that has been stained with a fluorescent probe for actin filaments. (A) The conventional, unprocessed image is blurred (more than...)
The confocal microscope has been used to resolve the structure of numerous complex three-dimensional objects (Figure 9-twenty), including the networks of cytoskeletal fibers in the cytoplasm and the arrangements of chromosomes and genes in the nucleus.

Effigy ix-20
Three-dimensional reconstruction from confocal microscope images. (A) Pollen grains, in this case from a passion flower, have a circuitous sculptured jail cell wall that contains fluorescent compounds. Images obtained at different depths through the grain, using (more...)
The relative merits of deconvolution methods and confocal microscopy for three-dimensional optical microscopy are withal the subject of contend. Confocal microscopes are generally easier to utilise than deconvolution systems and the final optical sections tin be seen quickly. On the other hand, modernistic, cooled CCD (charge-coupled device) cameras used for deconvolution systems are extremely efficient at collecting pocket-sized amounts of calorie-free, and they can be used to make detailed iii-dimensional images from specimens that are also weakly stained or as well easily damaged by brilliant calorie-free for confocal microscopy.
The Electron Microscope Resolves the Fine Structure of the Jail cell
The relationship betwixt the limit of resolution and the wavelength of the illuminating radiation (see Figure ix-six) holds true for any grade of radiations, whether it is a beam of lite or a beam of electrons. With electrons, however, the limit of resolution can be made very small. The wavelength of an electron decreases as its velocity increases. In an electron microscope with an accelerating voltage of 100,000 V, the wavelength of an electron is 0.004 nm. In theory the resolution of such a microscope should be near 0.002 nm, which is x,000 times that of the light microscope. Considering the aberrations of an electron lens are considerably harder to right than those of a glass lens, even so, the practical resolving ability of virtually modern electron microscopes is, at best, 0.1 nm (one Å) (Figure 9-21). This is because but the very center of the electron lenses can exist used, and the effective numerical aperture is tiny. Furthermore, problems of specimen preparation, contrast, and radiation damage have generally limited the normal constructive resolution for biological objects to ii nm (xx Å). This is nonetheless about 100 times improve than the resolution of the lite microscope. Moreover, in recent years, the performance of electron microscopes has been improved past the development of electron illumination sources called field emission guns. These very brilliant and coherent sources can substantially improve the resolution accomplished. The major landmarks in the development of electron microscopy are listed in Tabular array 9-2.

Figure 9-21
The limit of resolution of the electron microscope. This transmission electron micrograph of a sparse layer of aureate shows the individual files of atoms in the crystal as bright spots. The altitude between adjacent files of gilt atoms is well-nigh 0.two nm (2 (more than...)
Table 9-2
Major Events in the Development of the Electron Microscope and Its Application to Jail cell Biology.
In overall blueprint the transmission electron microscope (TEM) is similar to a calorie-free microscope, although information technology is much larger and upside down (Figure 9-22). The source of illumination is a filament or cathode that emits electrons at the tiptop of a cylindrical column most 2 thou high. Since electrons are scattered by collisions with air molecules, air must kickoff be pumped out of the cavalcade to create a vacuum. The electrons are then accelerated from the filament past a nearby anode and immune to laissez passer through a tiny hole to form an electron beam that travels down the cavalcade. Magnetic coils placed at intervals along the column focus the electron beam, just as glass lenses focus the calorie-free in a light microscope. The specimen is put into the vacuum, through an airlock, into the path of the electron axle. As in light microscopy, the specimen is usually stained—in this instance, with electron-dense material, every bit we encounter in the next section. Some of the electrons passing through the specimen are scattered past structures stained with the electron-dense material; the remainder are focused to course an image, in a manner analogous to the way an epitome is formed in a light microscope—either on a photographic plate or on a phosphorescent screen. Because the scattered electrons are lost from the beam, the dense regions of the specimen testify upward in the image equally areas of reduced electron flux, which look dark.

Figure 9-22
The primary features of a light microscope and a transmission electron microscope. These drawings emphasize the similarities of overall design. Whereas the lenses in the light microscope are made of glass, those in the electron microscope are magnetic (more than...)
Biological Specimens Require Special Preparation for the Electron Microscope
In the early days of its application to biological materials, the electron microscope revealed many previously unimagined structures in cells. But before these discoveries could be made, electron microscopists had to develop new procedures for embedding, cut, and staining tissues.
Since the specimen is exposed to a very high vacuum in the electron microscope, there is no possibility of viewing it in the living, wet state. Tissues are usually preserved by fixation—first with glutaraldehyde, which covalently cross-links protein molecules to their neighbors, and then with osmium tetroxide, which binds to and stabilizes lipid bilayers as well as proteins (Figure 9-23). Because electrons have very express penetrating power, the fixed tissues normally have to be cutting into extremely sparse sections (50–100 nm thick, about 1/200 the thickness of a unmarried cell) before they are viewed. This is achieved past dehydrating the specimen and permeating information technology with a monomeric resin that polymerizes to grade a solid block of plastic; the cake is and so cutting with a fine glass or diamond knife on a special microtome. These thin sections, gratis of water and other volatile solvents, are placed on a small circular metal filigree for viewing in the microscope (Figure 9-24).

Figure 9-23
Ii common chemic fixatives used for electron microscopy. The ii reactive aldehyde groups of glutaraldehyde enable it to cantankerous-link various types of molecules, forming covalent bonds between them. Osmium tetroxide is reduced past many organic compounds (more...)
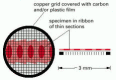
Effigy 9-24
The copper filigree that supports the thin sections of a specimen in a TEM.
The steps required to ready biological material for viewing in the electron microscope have challenged electron microscopists from the commencement. How can we be sure that the image of the fixed, dehydrated, resin-embedded specimen finally seen bears whatsoever relation to the delicate aqueous biological system that was originally present in the living jail cell? The best current approaches to this problem depend on rapid freezing. If an aqueous organization is cooled fast enough to a low plenty temperature, the water and other components in it do not have fourth dimension to rearrange themselves or crystallize into ice. Instead, the water is supercooled into a rigid but noncrystalline state—a "glass"—chosen vitreous water ice. This country tin be achieved by slamming the specimen onto a polished copper block cooled by liquid helium, by plunging it into or spraying it with a jet of a coolant such as liquid propane, or by cooling it at loftier pressure level.
Some frozen specimens can exist examined directly in the electron microscope using a special, cooled specimen holder. In other cases the frozen block tin be fractured to reveal interior surfaces, or the surrounding water ice tin be sublimed away to betrayal external surfaces. Withal, we often want to examine thin sections, and to accept them stained to yield acceptable contrast in the electron microscope paradigm (discussed farther below). A compromise is therefore to rapid-freeze the tissue, so replace the water, maintained in the vitreous (glassy) state, past organic solvents, and finally embed the tissue in plastic resin, cut sections, and stain. Although technically still difficult, this approach stabilizes and preserves the tissue in a condition very close to its original living state.
Contrast in the electron microscope depends on the atomic number of the atoms in the specimen: the higher the diminutive number, the more electrons are scattered and the greater the contrast. Biological tissues are composed of atoms of very low diminutive number (mainly carbon, oxygen, nitrogen, and hydrogen). To brand them visible, they are usually impregnated (before or after sectioning) with the salts of heavy metals such equally uranium and lead. Different cellular constituents are revealed with various degrees of contrast according to their degree of impregnation, or "staining," with these salts. Lipids, for example, tend to stain darkly after osmium fixation, revealing the location of cell membranes (Effigy 9-25).
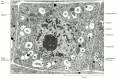
Figure ix-25
A root-tip prison cell stained with osmium and other heavy metal ions. The cell wall, nucleus, vacuoles, mitochondria, endoplasmic reticulum, Golgi apparatus, and ribosomes are hands visible in this transmission electron micrograph. (Courtesy of Brian Gunning.) (more...)
Specific Macromolecules Tin Be Localized by Immunogold Electron Microscopy
We have seen how antibodies can be used in conjunction with fluorescence microscopy to localize specific macromolecules. An analogous method—immunogold electron microscopy—can exist used in the electron microscope. The usual procedure is to incubate a sparse section with a specific primary antibody, and and so with a secondary antibiotic to which a colloidal gold particle has been fastened. The golden particle is electron-dense and can be seen every bit a black dot in the electron microscope (Figure 9-26).
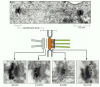
Effigy 9-26
Localizing proteins in the electron microscope. Immunogold electron microscopy is used hither to localize iv different poly peptide components to particular locations within the spindle pole body of yeast. At the top is a thin section of a yeast mitotic spindle (more...)
Sparse sections often fail to convey the iii-dimensional arrangement of cellular components in the TEM and can be very misleading: a linear structure such as a microtubule may appear in department as a pointlike object, for instance, and a section through protruding parts of a single irregularly shaped solid body may give the advent of two or more separate objects. The third dimension can be reconstructed from serial sections (Figure nine-27), but this is still a lengthy and wearisome process.

Figure 9-27
A three-dimensional reconstruction from series sections. Unmarried thin sections sometimes requite misleading impressions. In this example, well-nigh sections through a prison cell containing a branched mitochondrion seem to contain two or three dissever mitochondria. (more...)
Even thin sections, still, have a significant depth compared to the resolution of the electron microscope, so they can also exist misleading in an opposite way. The optical design of the electron microscope—the very pocket-sized discontinuity used—produces a large depth of field, so the image seen corresponds to a superimposition (a projection) of the structures at unlike depths. A further complication for immunogold labeling is that the antibodies and colloidal gold particles do not penetrate into the resin used for embedding; therefore, they only detect antigens right at the surface of the section. This means that commencement, the sensitivity of detection is low, since antigen molecules present in the deeper parts of the section are not detected, and second, ane may go a false impression of which structures contain the antigen and which exercise not. A solution to this trouble is to perform the labeling before embedding the specimen in plastic, when the cells and tissues are still fully accessible to labeling reagents. Extremely small gold particles, about i nm in diameter, work best for this procedure. Such small gold particles are usually not straight visible in the final sections, and so additional argent or gilded is nucleated around the 1 nm gold particles in a chemical process very much like photographic evolution.
Images of Surfaces Can Exist Obtained by Scanning Electron Microscopy
A scanning electron microscope (SEM) directly produces an image of the three-dimensional construction of the surface of a specimen. The SEM is usually a smaller, simpler, and cheaper device than a transmission electron microscope. Whereas the TEM uses the electrons that take passed through the specimen to form an image, the SEM uses electrons that are scattered or emitted from the specimen'due south surface. The specimen to be examined is fixed, dried, and coated with a sparse layer of heavy metal. Alternatively, information technology tin can exist rapidly frozen, and then transferred to a cooled specimen stage for straight examination in the microscope. Often an unabridged constitute or small animal can be put into the microscope with very picayune grooming (Effigy 9-28). The specimen, prepared in whatever of these means, is so scanned with a very narrow beam of electrons. The quantity of electrons scattered or emitted every bit this primary beam bombards each successive point of the metallic surface is measured and used to control the intensity of a second beam, which moves in synchrony with the primary beam and forms an image on a television screen. In this way, a highly enlarged paradigm of the surface every bit a whole is congenital up (Figure ix-29).

Figure 9-28
A developing wheat bloom, or spike. This delicate blossom spike was rapidly frozen, coated with a thin metallic picture, and examined in the frozen state in a SEM. This micrograph, which is at a depression magnification, demonstrates the big depth of focus of the (more...)
Figure 9-29
The scanning electron microscope. In a SEM, the specimen is scanned past a beam of electrons brought to a focus on the specimen by the electromagnetic coils that act as lenses. The quantity of electrons scattered or emitted equally the beam bombards each successive (more...)
The SEM technique provides smashing depth of field; moreover, since the amount of electron handful depends on the angle of the surface relative to the beam, the image has highlights and shadows that give it a iii-dimensional appearance (Figures ix-28 and 9-30). Only surface features can be examined, even so, and in most forms of SEM, the resolution attainable is non very high (most 10 nm, with an effective magnification of up to 20,000 times). As a result, the technique is usually used to study whole cells and tissues rather than subcellular organelles. Very high-resolution SEMs take, nevertheless, been recently adult with a brilliant coherent-field emission gun as the electron source. This type of SEM can produce images that rival TEM images in resolution (Figure 9-31).

Effigy 9-30
Scanning electron microscopy. (A) A scanning electron micrograph of the stereocilia projecting from a pilus cell in the inner ear of a bullfrog. For comparison, the same structure is shown past (B) differential-interference-contrast light microscopy and (more...)
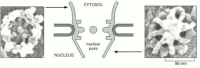
Figure nine-31
The nuclear pore. Chop-chop frozen nuclear envelopes were imaged in a high-resolution SEM, equipped with a field emission gun as the source of electrons. These views of each side of a nuclear pore correspond the limit of resolution of the SEM, and should (more...)
Metal Shadowing Allows Surface Features to Exist Examined at High Resolution by Manual Electron Microscopy
The TEM can as well exist used to study the surface of a specimen—and more often than not at a higher resolution than in the SEM—in such a style that individual macromolecules can be seen. Every bit in scanning electron microscopy, a sparse film of a heavy metal such as platinum is evaporated onto the dried specimen. The metal is sprayed from an oblique angle then as to deposit a coating that is thicker in some places than others—a process known as shadowing considering a shadow consequence is created that gives the image a three-dimensional appearance.
Some specimens coated in this fashion are sparse enough or pocket-size enough for the electron beam to penetrate them directly. This is the case for private molecules, viruses, and cell walls—all of which tin can be stale down, before shadowing, onto a flat supporting picture fabricated of a material that is relatively transparent to electrons, such as carbon or plastic. For thicker specimens, the organic material of the cell must exist dissolved away later on shadowing so that merely the sparse metal replica of the surface of the specimen is left. The replica is reinforced with a film of carbon then it can exist placed on a grid and examined in the transmission electron microscope in the ordinary way (Figure 9-32).

Effigy 9-32
The preparation of a metal-shadowed replica of the surface of a specimen. Note that the thickness of the metal reflects the surface contours of the original specimen.
Freeze-Fracture and Freeze-Compose Electron Microscopy Provide Views of Surfaces Inside the Cell
Freeze-fracture electron microscopy provides a way of visualizing the interior of cell membranes. Cells are frozen (as described above) and then the frozen block is croaky with a pocketknife blade. The fracture plane often passes through the hydrophobic middle of lipid bilayers, thereby exposing the interior of cell membranes. The resulting fracture faces are shadowed with platinum, the organic cloth is dissolved away, and the replicas are floated off and viewed in the electron microscope (see Effigy nine-32). Such replicas are studded with small-scale bumps, called intramembrane particles, which represent big transmembrane proteins. The technique provides a convenient and dramatic way to visualize the distribution of such proteins in the aeroplane of a membrane (Figure 9-33).

Figure 9-33
The thylakoid membranes from the chloroplast of a plant cell. In this freeze-fracture electron micrograph, the thylakoid membranes, which perform photosynthesis, are stacked up in multiple layers (see Figure 14-34). The plane of the fracture has moved (more...)
Another related replica method is freeze-compose electron microscopy, which can be used to examine either the outside or interior of cells. In this technique, the frozen block is cracked with a pocketknife bract as described above. But now the water ice level is lowered effectually the cells (and to a lesser extent within the cells) by the sublimation of water ice in a vacuum as the temperature is raised—a procedure called freeze-drying. The parts of the cell exposed by this etching process are and then shadowed as before to make a platinum replica. This technique exposes structures in the interior of the cell and can reveal their three-dimensional organization with exceptional clarity (Figure 9-34).

Figure nine-34
A regular array of poly peptide filaments in an insect muscle. To obtain this image, the musculus cells were rapidly frozen to liquid helium temperature, fractured through the cytoplasm, and subjected to deep etching. A metal-adumbral replica was then prepared (more than...)
Negative Staining and Cryoelectron Microscopy Let Macromolecules to Be Viewed at High Resolution
Although isolated macromolecules, such as DNA or large proteins, tin can be visualized readily in the electron microscope if they are adumbral with a heavy metal to provide dissimilarity, finer item tin can exist seen by using negative staining. In this technique, the molecules, supported on a thin moving-picture show of carbon, are washed with a concentrated solution of a heavy-metal table salt such as uranyl acetate. After the sample has dried, a very thin film of metal common salt covers the carbon pic everywhere except where it has been excluded past the presence of an adsorbed macromolecule. Considering the macromolecule allows electrons to pass much more readily than does the surrounding heavy-metallic stain, a reversed or negative image of the molecule is created. Negative staining is specially useful for viewing large macromolecular aggregates such as viruses or ribosomes, and for seeing the subunit structure of protein filaments (Effigy 9-35).

Figure 9-35
Negatively stained actin filaments. In this transmission electron micrograph, each filament is about 8 nm in diameter and is seen, on shut inspection, to exist equanimous of a helical concatenation of globular actin molecules. (Courtesy of Roger Craig.)
Shadowing and negative staining tin provide loftier-contrast surface views of small macromolecular assemblies, but both techniques are limited in resolution by the size of the smallest metallic particles in the shadow or stain used. Recent methods provide an alternative that has immune even the interior features of iii-dimensional structures such as viruses to be visualized directly at loftier resolution. In this technique, chosen cryoelectron microscopy, rapid freezing to form vitreous water ice is again the fundamental. A very thin (most 100 nm) film of an aqueous interruption of virus or purified macromolecular circuitous is prepared on a microscope grid. The specimen is then rapidly frozen by plunging information technology into a coolant. A special sample holder is used to go along this hydrated specimen at -160°C in the vacuum of the microscope, where information technology tin can exist viewed directly without fixation, staining, or drying. Unlike negative staining, in which what is seen is the envelope of stain exclusion around the particle, hydrated cryoelectron microscopy produces an image from the macromolecular construction itself. However, to extract the maximum corporeality of structural information, special epitome-processing techniques must be used, as we describe next.
Multiple Images Can Exist Combined to Increase Resolution
Whatever image, whether produced by an electron microscope or by an optical microscope, is made by particles—electrons or photons—striking a detector of some sort. But these particles are governed past breakthrough mechanics, and then the numbers reaching the detector are anticipated merely in a statistical sense. In the limit of very large numbers of particles, the distribution at the detector is accurately adamant by the imaged specimen. Notwithstanding, with smaller numbers of particles, this underlying structure in the image is obscured past the statistical fluctuations in the numbers of particles detected in each region. Random variability that confuses the underlying image of the specimen itself is referred to as noise. Noise is a particularly astringent trouble for electron microscopy of unstained macromolecules, just it is also of import in light microscopy at depression light levels. A protein molecule can tolerate a dose of but a few tens of electrons per foursquare nanometer without damage, and this dose is orders of magnitude beneath what is needed to define an image at atomic resolution.
The solution is to obtain images of many identical molecules—perhaps tens of thousands of private images—and combine them to produce an averaged prototype, revealing structural details that were hidden by the racket in the original images. Before the individual images tin be combined, notwithstanding, they must be aligned with each other. Sometimes it is possible to induce proteins and complexes to form crystalline arrays, in which each molecule is held in the aforementioned orientation in a regular lattice. In this case, the alignment trouble is easily solved, and several protein structures have been determined at atomic resolution by this blazon of electron crystallography. In principle, withal, crystalline arrays are non admittedly required. With the assistance of a computer, the images of randomly distributed molecules can be processed and combined to yield high-resolution reconstructions, as we at present explain.
Views from Unlike Directions Can Be Combined to Give Three-dimensional Reconstructions
The detectors used to record images from electron microscopes produce ii-dimensional pictures. Considering of the large depth of field of the microscope, all the parts of the three-dimensional specimen are in focus, and the resulting paradigm is a projection of the structure forth the viewing direction. The lost data in the third dimension can be recovered if nosotros have views of the same specimen from many unlike directions. The computational methods for this technique were worked out in the 1960s, and they are widely used in medical computed tomography (CT) scans. In a CT scan, the imaging equipment is moved around the patient to generate the different views. In electron-microscope (EM) tomography, the specimen holder is tilted in the microscope, which achieves the same result. In this fashion, one can make it at a three-dimensional reconstruction, in a called standard orientation, by combining a set of views of many identical molecules in the microscope'southward field of view. Each view will be individually very noisy, only by combining them in three dimensions and taking an boilerplate, the noise tin can exist largely eliminated, yielding a clear view of the molecular structure.
EM tomography is now widely applied for determining both molecular structures, using either crystalline or noncrystalline specimens, and larger objects such as thin sections of cells and organelles. It is a particularly successful technique for structures that accept some intrinsic symmetry, such as helical or icosahedral viruses, because it makes the task of alignment easier and more accurate. Figure 9-36 shows the structure of an icosahedral virus that has been adamant at high resolution by the combination of many particles and multiple views, and Effigy nine-37 shows the structure of a ribosome determined in the same style.

Figure 9-36
EM tomography. Spherical poly peptide shells of the hepatitis B virus are preserved in a thin film of ice (A) and imaged in the manual electron microscope. Thousands of individual particles were combined past EM tomography to produce the 3-dimensional (more...)
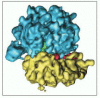
Figure 9-37
The 3-dimensional structure of the 70S ribosome from Due east. coli determined by EM tomography. The small subunit is colored xanthous, the big subunit blue. The overall resolution is 11.2 Å. (From I.S. Gabashvili et al., Jail cell 100: 537–549, (more...)
With crystalline arrays, a resolution of 0.3 nm has been achieved past electron microscopy—enough to begin to see the internal atomic arrangements in a protein and to rival x-ray crystallography in resolution. With single-particle reconstruction, the limit at the moment is almost 0.viii nm, enough to identify protein subunits and domains, and limited poly peptide secondary structure. Although electron microscopy is unlikely to supersede ten-ray crystallography (discussed in Chapter viii) as a method for macromolecular structure conclusion, it has some very clear advantages. First, it does not absolutely require crystalline specimens. Second, information technology can bargain with extremely big complexes—structures that may be besides large or too variable to crystallize satisfactorily. Electron microscopy provides a span between the scale of the single molecule and that of the whole cell.
Summary
Many lite-microscope techniques are available for observing cells. Cells that have been fixed and stained can be studied in a conventional light microscope, while antibodies coupled to fluorescent dyes can exist used to locate specific molecules in cells in a fluorescence microscope. Living cells can be seen with phase-contrast, differential-interference-contrast, dark-field, or bright-field microscopes. All forms of light microscopy are facilitated past electronic image-processing techniques, which raise sensitivity and refine the image. Confocal microscopy and epitome deconvolution both provide thin optical sections and tin can be used to reconstruct three-dimensional images.
Determining the detailed construction of the membranes and organelles in cells requires the college resolution attainable in a manual electron microscope. Specific macromolecules can exist localized with colloidal aureate linked to antibodies. Three-dimensional views of the surfaces of cells and tissues are obtained by scanning electron microscopy. The shapes of isolated macromolecules that have been adumbral with a heavy metal or outlined by negative staining can also be readily determined by electron microscopy. Using computational methods, multiple images and views from different directions are combined to produce detailed reconstructions of macromolecules and molecular complexes through a technique known as electron-microscope tomography.

Electron Micrograph Of Cell Labeled,
Source: https://www.ncbi.nlm.nih.gov/books/NBK26880/
Posted by: richardswalouteemper.blogspot.com
0 Response to "Electron Micrograph Of Cell Labeled"
Post a Comment